Le système nerveux et le système endocrinien permettent le maintient de l’homéostasie. Le système endocrinien sécrète des hormones dans le sang, leurs actions est lente mais soutenue dans le temps (cf. cours sur l’endocrinologie). Le système nerveux quant à lui permet la formation d’influx nerveux qui ont une action rapide mais brève, on parle ici de neuromédiateurs qui agissent sur de très courte distance (quelques µm) au niveau des synapses.
Le système nerveux est divisé en deux grandes zones : le système nerveux central (SNC) et le système nerveux périphérique (SNP). Le système nerveux central est constitué de l’encéphale (cerveau) et de la moelle épinière, et le système nerveux périphérique est constitué des ganglions nerveux et des nerfs : 12 paires de nerfs crâniens et 31 paires de nerfs rachidiens.
Le système nerveux a 3 fonctions essentielles :
-
Une fonction sensitive de détection grâce à des récepteurs qui détectent toutes les modifications de l’organisme et l’environnement extérieur.
-
Une fonction d’intégration et d’analyse des informations qu’il reçoit des récepteurs.
-
Une fonction motrice permettant la contraction des diverses cellules musculaires de l’organisme.
Toutes les informations de l’organisme affluent vers le SNC à partir de détecteurs sensoriels de différents types.
Le SNP est constitué de deux voies :
-
La voie sensitive (voie afférente) constituée de neurones sensitifs somatiques et viscéraux, et au niveau de laquelle la propagation des influx vient des récepteurs périphériques.
-
La voie motrice (voie efférente) constituée de neurones moteurs dont l’origine des influx est le SNC. Cette voie motrice peut elle-même être divisée en deux types de système nerveux :
-
Le système nerveux autonome (SNA), ou système nerveux végétatif (SNV), est involontaire. L’influx nerveux provenant du SNC est envoyé vers les muscles lisses, le myocarde et les glandes. Il possède le système sympathique (Σ) qui tend à activer les organes et le système parasympathique (pΣ) qui tend à les mettre au repos. Attention, les deux peuvent pourtant être excitateur et inhibiteur.
-
Le système nerveux somatique (SNS) est volontaire et l’influx nerveux provenant du SNC est envoyé vers les muscles striés squelettiques.
-
Les cellules nerveuses (neurones) sont les unités fonctionnelles du SNC et forment un réseau qui s’étend dans tout l’organisme. Bien qu’elles ne soient pas les plus nombreuses dans le système nerveux, ce sont les plus importantes. Pour indication le système nerveux possède 10% de neurones pour 90% de cellules gliales (leur rôle dans le système nerveux sera énoncé dans la suite du cours).
Ce sont des cellules post-mitotique (pour la majorité, étant hautement spécialisés) et excitable. En effet on sait aujourd’hui que certains neurones peuvent être produits au niveau de l’hippocampe. Leur excitabilité est due à un changement d’état très rapide qui est déterminé par un facteur extérieur. Les cellules nerveuses peuvent modifier leur anatomie et possèdent une grande longévité. Elles sont très sensibles à l’hypoglycémie et à l’hypoxie. En effet elles consomment presque exclusivement du glucose qui leurs est fournit par les cellules gliales. Les neurones sont indépendants les uns des autres, n’établissant que des contacts fonctionnels spécifiques appelés synapses. Ce sont également des cellules sécrétrices particulières qui peuvent avoir comme produit de sécrétion des neuromédiateurs, des neuromodulateurs ainsi que des neurohormones (GnRH).
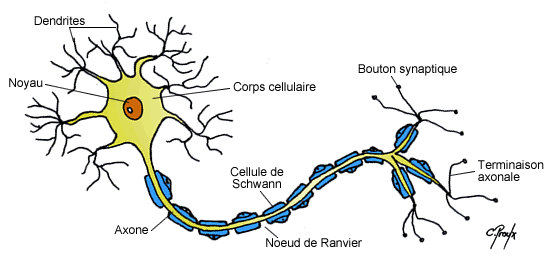
a) Morphologie du neurone
La morphologie du neurone est caractérisée par différentes structures :
-
Le corps cellulaire (ou soma ou péricaryon) : est la partie vitale de la cellule. Il est constitué d’un noyau qui détermine la forme du corps cellulaire, le cytoplasme y étant presque accolé, et dont le nucléole est volumineux reflétant ainsi la forte activité de synthèse de ces cellules. Le corps cellulaire est la portion centrale du neurone d’où émergent l’axone et les dendrites. Il possède les mêmes organites que dans la majorité des cellules ; on note cependant la présence d’amas de réticulum endoplasmique granuleux (lieu de synthèse protéique) que l’on appelle corps de Nissl, ainsi qu’une grande quantité de protéines du cytosquelette, appelées neurofibrilles, qui sont responsables de la communication intracellulaire. Un des rôles principaux du corps cellulaire est de synthétiser une grande partie des constituants nécessaires à la structure et aux fonctions du neurone.
L’axone : est un prolongement unique, fin, homogène, relativement linéaire et pouvant s’arboriser par la suite au niveau des nœuds de Ranvier. Il prend naissance au niveau d’une expansion conique du corps cellulaire appelée cône d’implantation (ou cône d’émergence) qui est également le lieu d’où partira le potentiel d’action (cf. suite du cours). L’axone peut se diviser en une ou plusieurs collatérales qui se termineront généralement par une arborisation terminale dont chaque extrémité, renflée, établit des contacts synaptiques avec les cellules cibles. Les neurones sont principalement constitués de neurofibrilles et de mitochondries qui fournissent l’énergie nécessaire aux mouvements des messagers intracellulaires et à la libération des vésicules synaptiques au niveau des extrémités axonales, appelées boutons synaptiques. L’axone est également le lieu de transports qui sont soit antérograde (vers les boutons synaptique) soit rétrograde (vers le corps cellulaire). Ce transport continuel représente un flux nécessaire à l’apport des différentes macromolécules tout au long de l’axone ; en effet les axones ne présentent aucunes structures responsables de la synthèse de protéines.
b) La gaine de myéline
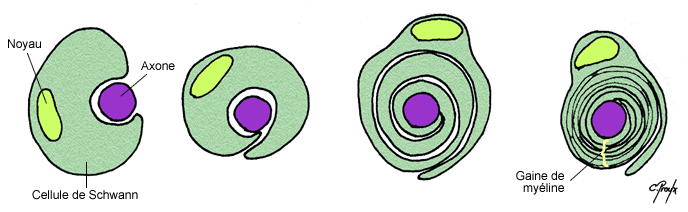
Les axones peuvent être recouverts par une gaine de myéline qui correspond à l’enroulement de couches phospholipidiques concentriques de manière discontinue sur l’ensemble de l’axone. En effet ces gaines sont espacées tous les 1 à 2 mm par les nœuds de Ranvier qui sont du coup amyéliniques et d’où peuvent émerger les collatérales de l’axone. Ces gaines sont formées à partir de 2 types cellulaires suivant si l’on se trouve dans le SNC ou le SNP :
-
Dans le SNC on trouve des oligodendrocytes qui envoient des prolongements de leurs corps cellulaires qui recouvreront les axones. Il peut ainsi participer à des gaines de neurones différents.
-
Dans le SNP on trouve des cellules de Schwann qui vont s’enrouler entièrement autour d’un axone unique. Leur noyau est renvoyé au niveau de la face la plus externe.
-
- Les neurones bipolaires (sensitif)
Les neurones multipolaires (moteur et sensitif)
De manière fonctionnelle il existe :
-
Les neurones sensoriels
-
Les neurones moteurs
-
Les inter-neurones, ce sont les plus nombreux et ils servent de lien entre les neurones dans le SNC.
Les cellules de la névroglie sont les cellules majoritaires du système nerveux. Elles sont étroitement liées aux neurones, de taille inférieure à ces derniers et ne forment aucune synapse chimique. Contrairement aux neurones, les cellules gliales peuvent se reproduire par mitose. Elles ont différents rôles au sein des tissus nerveux : l’isolement des tissus nerveux (cf. Oligodendrocytes et cellules de Schwann), les fonctions métaboliques (cf. Astrocytes), le soutien structural et une protection immunitaire (cf. Microglie). La macroglie correspond aux astrocytes et aux oligodendrocytes.
Les cellules gliales, toutes situées dans le SNC, sont de différents types :
-
Astrocytes : cellules de petite taille et de forme étoilée, dont leurs extrémités se finissent pas un renflement appelé pied astrocytaire. On distingue deux types d’astrocytes :
-
Les astrocytes de type I sont accolés aux capillaires pour en prélever des nutriments comme le glucose et le calcium et pour en déverser le potassium extracellulaire excédentaire. Il y a ainsi un contrôle de l’environnement ionique immédiat des neurones. Ces astrocytes participent, avec les cellules endothéliales, à la formation de la barrière hémato-encéphalique.
-
Les astrocytes de type II entourent les neurones et la fente synaptique (recapture du neurotransmetteur).
-
-
Oligodendrocytes : plus petits que les astrocytes. Ils possèdent un cytoplasme plus dense et sont présent dans la substance blanche. Les oligodendrocytes ont comme principale fonction d’envelopper les axones des neurones formant ainsi les gaines de myélines qui donneront la couleur blanche de la substance blanche. Ils peuvent être comparés aux cellules de Schwann du SNP. Un seul oligodendrocyte pourra former plusieurs gaines de myéline sur plusieurs neurones différents (contrairement aux cellules de Schwann).
-
Microglie : cellules de petite taille, représentant 5-20% de la population gliale totale. Les cellules microgliales sont présentes en plus grande quantité dans la substance grise que dans la blanche. Elles sont activées suite à une atteinte du SNC puis se déplacent vers les sites atteints où elles se multiplient. Ces cellules appartiennent au groupe des macrophages et phagocytent donc les cellules mortes et les corps étrangers.
-
Cellules épendymaires : ce sont des cellules épithéliales (épithélium simple) situées dans certaines cavités de l’encéphale que l’on appelle ventricules. Elles y fabriquent le liquide céphalorachidien qui protège l’encéphale et la moelle épinière tout en participant à satisfaire l’ensemble des besoins physiologiques des cellules du tissu nerveux. Les cellules épendymaires spécialisées qui fabriquent le liquide céphalo-rachidien forment les structures anatomiques appelées les plexus choroïdes. On retrouve aussi ces cellules dans le canal de l’épendyme situé à l’intérieur de la moelle épinière.
1) Potentiels et influx nerveux
L’influx nerveux est le potentiel électrique se déplaçant sur l’axone après que le neurone ait été stimulé. L’excitabilité est la capacité à réagir à un stimulus et à le convertir en influx nerveux. La conductivité est la capacité de propagation et de transmission de l’influx nerveux.
La transmission de l’influx nerveux se fait des dendrites jusqu’à l’axone. En effet l’arbre somato-dendritique représente le pôle récepteur du neurone et l’axone (ou collatérales) représente le pôle émetteur du neurone. Attention cela ne veut pas dire que l’axone ne peut pas jouer le rôle de récepteur.
La communication entre neurones se fait grâce :
-
aux potentiels d’action conduit au niveau des axones sur de longues distances avec peu de pertes.
-
aux potentiels gradués conduit au niveau des dendrites sur de courtes distances avec des pertes importantes.
Les neurones, comme toutes les cellules de l’organisme, sont soumise à une différence de potentiel membranaire (ddp) due aux différences de concentration ioniques de part et d’autre de la membrane. Du côté extracellulaire ce sont surtout les ions Na et Cl- qui sont présents, et du côté intracellulaire ce sont surtout les ions K et les protéines qui sont présents.
On note que les ions K sont ceux qui possèdent la plus grande conductance au sein de la membrane (5 fois plus élevé que les autres ions), il attire donc le potentiel de membrane vers son potentiel d’équilibre (-80 mV) donné par l’équation de Nernst. Le gradient de concentration des ions potassique les pousse à sortir de la cellule, mais l’existence de charge positive dans le milieu extracellulaire créé un gradient électrique de sens contraire au gradient de concentration des ions K . Autrement dit le potentiel de repos est atteint à l’équilibre, lorsque les forces dues au gradient électrique (qui poussent à faire rentrer les ions K dans la cellule) sont égales aux forces dues au gradient de concentration (qui poussent à faire sortir les ions K de la cellule).
On arrive à un équilibre des forces, la différence de potentiel est alors de -70 mV. Elle se maintient même si Na parvient à rentrer dans la cellule, et ceci par régulation des pompes Na /K . On peut faire la remarque que le potentiel de membrane est nul lorsque la concentration en ions chargés négativement est égale à la concentration en ions chargés positivement, et ce dans le milieu intracellulaire et extracellulaire.
b) Le potentiel gradué
Le potentiel gradué est une inversion locale et de courte durée du potentiel membranaire. Il apparaît au niveau des dendrites et des corps cellulaires et est déclenché par une stimulation extérieure à la cellule (inversion locale de la polarité membranaire). Suite à cette stimulation il y a apparition d’un courant électrique local qui se propageant bilatéralement par rapport au point de stimulation et dont l’intensité diminue avec la distance.
Il est dit gradué, car son voltage est proportionnel à l’intensité de la stimulation. Ce potentiel gradué arrivera jusqu’au corps cellulaire et si son voltage est suffisant il y aura formation d’un potentiel d’action.
c) Le potentiel d’action
Le potentiel d’action est une variation transitoire du potentiel membranaire déclenchée suite à une stimulation, formée au niveau du cône d’émergence et dont la propagation est axonique, unidirectionnelle, avec une intensité qui ne diminue pas avec la distance.
La stimulation peut provenir d’un autre neurone, ou de la stimulation d’un récepteur sensitif qui peut être présent à la surface de l’organisme (peau) ou bien même dans l’organisme lui-même (au niveau des organes). Il peut également y avoir des potentiels d’action auto-entretenu, c’est le cas du cœur (cf. cours physiologie du système cardiovasculaire). Si cette stimulation est suffisante, c’est-à-dire si elle dépasse le seuil de déclenchement du neurone, alors il y aura création du potentiel d’action. Attention le seuil n’est pas le même pour tous les neurones.
-
La dépolarisation correspond à une augmentation de la perméabilité sodique, qui entraîne une réduction du potentiel membranaire. L’intérieur de la membrane est moins négatif et le potentiel s’approche de 0.
-
La repolarisation rapide du point dépolarisé correspond à la fermeture des canaux sodiques et à l’ouverture des canaux potassique plus ou moins décalé dans le temps.
-
L’hyperpolarisation correspond à une sortie en excès d’ions K lors de la repolarisation ce qui entraîne une augmentation de la différence de potentiel membranaire, plus importante que la différence de potentiel présente au repos. Il y a dès lors intervention des pompes Na /K pour rétablir les concentrations ioniques.

Dans le cas des neurones amyéliniques, le potentiel d’action possède la même amplitude tout du long ; la dépolarisation en un point induit la dépolarisation du point voisin, la propagation est lente. Il existe une période réfractaire. Le potentiel d’action se déplace en sens unique du cône d’émergence vers les terminaisons.
Dans le cas des neurones myélinisés, il y a création successive des potentiels d’action le long de l’axone ; ceux-ci vont s’éloigner du site d’excitation initiale. La vitesse de transmission dépend du diamètre de la fibre (s’il augmente, la vitesse augmente). La conduction est dite conduction saltatoire (de nœud en nœud) et la propagation est rapide. Après la repolarisation, la membrane demeure inerte un certain temps ; les canaux à Na ne peuvent pas s’ouvrir (période réfractaire). Au niveau des nœuds de Ranvier on met en évidence une grande concentration de canaux sodique voltage dépendant qui sont responsable de la propagation du potentiel d’action.
-
Remarques :Les invertébrés n’ont pas de neurones myélinisés ; ils doivent donc compenser par leur diamètre (le plus gros axone connu est l’axone géant du réflexe de fuite du calamar ; v=20 m.s-1, diamètre=1mm). Une fibre non myélinisée devrait avoir un calibre de plusieurs centimètres pour conduire l’influx à la même vitesse (100 m/s) qu’une fibre myélinisée de 20 micromètres de diamètre.
La synapse correspond au point de connexion fonctionnel existant entre deux neurones. Un millimètre cube de substance grise du cortex peut contenir 5 millions de synapses. Les synapses peuvent être électriques ou chimiques :
Les synapses chimiques, quant à elles, sont présentent uniquement dans le tissu nerveux et ce sont elles que nous allons expliquer dans la suite de ce cours. Suivant les cellules impliquées on aura des :
-
Synapses neuro-neuronales : jonction entre 2 neurones. Parmi elles on trouve les synapses axo-dendritiques (entre l’axone et les dendrites) et axo-somatiques (entre l’axone et le corps cellulaires).
-
Synapses neuro-effectrices : jonction entre un neurone (moteur) et une cellule effectrice (cellule musculaire ou cellule sécrétrice d’une glande, comme la glande surrénale par exemple).
-
Synapses sensori-neuronales : jonction entre des cellules sensorielles et des neurones.
- Il peut être excitateur grâce à l’entrée d’ions sodium Na , on parle de potentiel post-synaptique excitateur (PPSE). Le PPSE n’est créé que si la dépolarisation dépasse le seuil permettant la formation d’un potentiel d’action et donc la propagation de l’influx nerveux.
Il peut être inhibiteur, on parle de potentiel post-synaptique inhibiteur (PPSI). Le PPSI est créé par l’entré d’ion chlorure Cl- ou la sortie d’ion potassique K qui permettent une hyperpolarisation diminuant l’excitabilité neuronale.
Le potentiel d’action est soumis à la loi du tout ou rien, c’est-à-dire que quelque soit le courant créé par le stimulus, s’il arrive jusqu’au seuil du neurone il permettra une dépolarisation complète, mais s’il n’arrive pas jusqu’au seuil, il ne se passera rien du tout :
-
Si la dépolarisation ne passe pas le seuil (-50mV / -40mV), il n’y aura pas d’influx.
-
Si la dépolarisation passe le seuil, la dépolarisation se poursuit jusqu’à + 40mV.
-
Quelque soit l’intensité du stimulus, la dépolarisation ne dépasse pas les + 40mV.
synapse.
La période réfractaire est la période durant laquelle l’axone ne pourra plus propager de potentiel d’action après un premier potentiel d’action. Il y a présence de deux types de périodes réfractaires suivant l’avancé du premier potentiel d’action.
-
Pendant la période réfractaire absolue le neurone ne peut dans aucun cas répondre à une nouvelle stimulation. Elle est à prendre en compte jusqu’à ce que le potentiel, lors de la repolarisation, revienne jusqu’au potentiel seuil.
-
Pendant la période réfractaire relative le neurone peut répondre à une nouvelle stimulation, mais elle nécessite un stimulus d’autant plus grand qu’on se situe proche de la période réfractaire absolue. Elle prend place tout de suite après la période réfractaire absolue et jusqu’à ce que le potentiel soit de nouveau au niveau du potentiel de membrane de repos.
