Salut à tout le monde
Aujourd’hui, Je vous ramène
l’examen de système d’exploitation année 2022-2010
de l’université IBN KHALDOUN TIARET (Mr Bekki )

Salut à tout le monde
Aujourd’hui, Je vous ramène
l’examen de système d’exploitation année 2022-2010
de l’université IBN KHALDOUN TIARET (Mr Bekki )
</object
En cliquant sur les flacons une goutte de sérum, de globules ou de sang est déposée dans les cavités de la plaque à réactions. Chaque cavité correspond à un produit test, le sang à tester étant réparti dans toutes les cavités.
Remarque : l’image ci-dessous représente une
agglutination.

M-M LEPAGE & R MONTANARI
[web]http://c.coupin.free.fr/cartable/troisieme/protection%20_organisme/gpesang/menu.htm[/web]
.
Le système nerveux et le système endocrinien permettent le maintient de l’homéostasie. Le système endocrinien sécrète des hormones dans le sang, leurs actions est lente mais soutenue dans le temps (cf. cours sur l’endocrinologie). Le système nerveux quant à lui permet la formation d’influx nerveux qui ont une action rapide mais brève, on parle ici de neuromédiateurs qui agissent sur de très courte distance (quelques µm) au niveau des synapses.
Le système nerveux est divisé en deux grandes zones : le système nerveux central (SNC) et le système nerveux périphérique (SNP). Le système nerveux central est constitué de l’encéphale (cerveau) et de la moelle épinière, et le système nerveux périphérique est constitué des ganglions nerveux et des nerfs : 12 paires de nerfs crâniens et 31 paires de nerfs rachidiens.
Le système nerveux a 3 fonctions essentielles :
Toutes les informations de l’organisme affluent vers le SNC à partir de détecteurs sensoriels de différents types.
Le SNP est constitué de deux voies :
Les cellules nerveuses (neurones) sont les unités fonctionnelles du SNC et forment un réseau qui s’étend dans tout l’organisme. Bien qu’elles ne soient pas les plus nombreuses dans le système nerveux, ce sont les plus importantes. Pour indication le système nerveux possède 10% de neurones pour 90% de cellules gliales (leur rôle dans le système nerveux sera énoncé dans la suite du cours).
Ce sont des cellules post-mitotique (pour la majorité, étant hautement spécialisés) et excitable. En effet on sait aujourd’hui que certains neurones peuvent être produits au niveau de l’hippocampe. Leur excitabilité est due à un changement d’état très rapide qui est déterminé par un facteur extérieur. Les cellules nerveuses peuvent modifier leur anatomie et possèdent une grande longévité. Elles sont très sensibles à l’hypoglycémie et à l’hypoxie. En effet elles consomment presque exclusivement du glucose qui leurs est fournit par les cellules gliales. Les neurones sont indépendants les uns des autres, n’établissant que des contacts fonctionnels spécifiques appelés synapses. Ce sont également des cellules sécrétrices particulières qui peuvent avoir comme produit de sécrétion des neuromédiateurs, des neuromodulateurs ainsi que des neurohormones (GnRH).
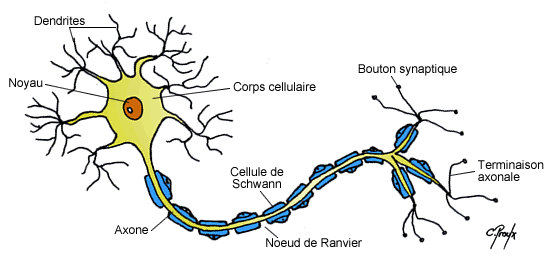
a) Morphologie du neurone
La morphologie du neurone est caractérisée par différentes structures :
L’axone : est un prolongement unique, fin, homogène, relativement linéaire et pouvant s’arboriser par la suite au niveau des nœuds de Ranvier. Il prend naissance au niveau d’une expansion conique du corps cellulaire appelée cône d’implantation (ou cône d’émergence) qui est également le lieu d’où partira le potentiel d’action (cf. suite du cours). L’axone peut se diviser en une ou plusieurs collatérales qui se termineront généralement par une arborisation terminale dont chaque extrémité, renflée, établit des contacts synaptiques avec les cellules cibles. Les neurones sont principalement constitués de neurofibrilles et de mitochondries qui fournissent l’énergie nécessaire aux mouvements des messagers intracellulaires et à la libération des vésicules synaptiques au niveau des extrémités axonales, appelées boutons synaptiques. L’axone est également le lieu de transports qui sont soit antérograde (vers les boutons synaptique) soit rétrograde (vers le corps cellulaire). Ce transport continuel représente un flux nécessaire à l’apport des différentes macromolécules tout au long de l’axone ; en effet les axones ne présentent aucunes structures responsables de la synthèse de protéines.
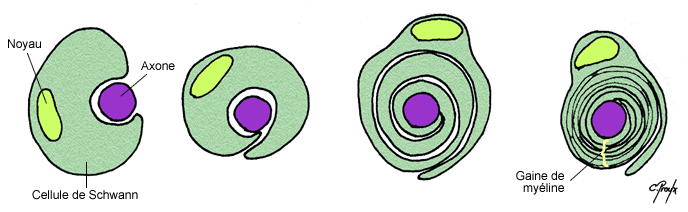
b) La gaine de myéline
Les axones peuvent être recouverts par une gaine de myéline qui correspond à l’enroulement de couches phospholipidiques concentriques de manière discontinue sur l’ensemble de l’axone. En effet ces gaines sont espacées tous les 1 à 2 mm par les nœuds de Ranvier qui sont du coup amyéliniques et d’où peuvent émerger les collatérales de l’axone. Ces gaines sont formées à partir de 2 types cellulaires suivant si l’on se trouve dans le SNC ou le SNP :
Les neurones multipolaires (moteur et sensitif)
De manière fonctionnelle il existe :
Les cellules de la névroglie sont les cellules majoritaires du système nerveux. Elles sont étroitement liées aux neurones, de taille inférieure à ces derniers et ne forment aucune synapse chimique. Contrairement aux neurones, les cellules gliales peuvent se reproduire par mitose. Elles ont différents rôles au sein des tissus nerveux : l’isolement des tissus nerveux (cf. Oligodendrocytes et cellules de Schwann), les fonctions métaboliques (cf. Astrocytes), le soutien structural et une protection immunitaire (cf. Microglie). La macroglie correspond aux astrocytes et aux oligodendrocytes.
Les cellules gliales, toutes situées dans le SNC, sont de différents types :
1) Potentiels et influx nerveux
L’influx nerveux est le potentiel électrique se déplaçant sur l’axone après que le neurone ait été stimulé. L’excitabilité est la capacité à réagir à un stimulus et à le convertir en influx nerveux. La conductivité est la capacité de propagation et de transmission de l’influx nerveux.
La transmission de l’influx nerveux se fait des dendrites jusqu’à l’axone. En effet l’arbre somato-dendritique représente le pôle récepteur du neurone et l’axone (ou collatérales) représente le pôle émetteur du neurone. Attention cela ne veut pas dire que l’axone ne peut pas jouer le rôle de récepteur.
La communication entre neurones se fait grâce :
Les neurones, comme toutes les cellules de l’organisme, sont soumise à une différence de potentiel membranaire (ddp) due aux différences de concentration ioniques de part et d’autre de la membrane. Du côté extracellulaire ce sont surtout les ions Na et Cl- qui sont présents, et du côté intracellulaire ce sont surtout les ions K et les protéines qui sont présents.
On note que les ions K sont ceux qui possèdent la plus grande conductance au sein de la membrane (5 fois plus élevé que les autres ions), il attire donc le potentiel de membrane vers son potentiel d’équilibre (-80 mV) donné par l’équation de Nernst. Le gradient de concentration des ions potassique les pousse à sortir de la cellule, mais l’existence de charge positive dans le milieu extracellulaire créé un gradient électrique de sens contraire au gradient de concentration des ions K . Autrement dit le potentiel de repos est atteint à l’équilibre, lorsque les forces dues au gradient électrique (qui poussent à faire rentrer les ions K dans la cellule) sont égales aux forces dues au gradient de concentration (qui poussent à faire sortir les ions K de la cellule).
On arrive à un équilibre des forces, la différence de potentiel est alors de -70 mV. Elle se maintient même si Na parvient à rentrer dans la cellule, et ceci par régulation des pompes Na /K . On peut faire la remarque que le potentiel de membrane est nul lorsque la concentration en ions chargés négativement est égale à la concentration en ions chargés positivement, et ce dans le milieu intracellulaire et extracellulaire.
b) Le potentiel gradué
Le potentiel gradué est une inversion locale et de courte durée du potentiel membranaire. Il apparaît au niveau des dendrites et des corps cellulaires et est déclenché par une stimulation extérieure à la cellule (inversion locale de la polarité membranaire). Suite à cette stimulation il y a apparition d’un courant électrique local qui se propageant bilatéralement par rapport au point de stimulation et dont l’intensité diminue avec la distance.
Il est dit gradué, car son voltage est proportionnel à l’intensité de la stimulation. Ce potentiel gradué arrivera jusqu’au corps cellulaire et si son voltage est suffisant il y aura formation d’un potentiel d’action.
c) Le potentiel d’action
Le potentiel d’action est une variation transitoire du potentiel membranaire déclenchée suite à une stimulation, formée au niveau du cône d’émergence et dont la propagation est axonique, unidirectionnelle, avec une intensité qui ne diminue pas avec la distance.
La stimulation peut provenir d’un autre neurone, ou de la stimulation d’un récepteur sensitif qui peut être présent à la surface de l’organisme (peau) ou bien même dans l’organisme lui-même (au niveau des organes). Il peut également y avoir des potentiels d’action auto-entretenu, c’est le cas du cœur (cf. cours physiologie du système cardiovasculaire). Si cette stimulation est suffisante, c’est-à-dire si elle dépasse le seuil de déclenchement du neurone, alors il y aura création du potentiel d’action. Attention le seuil n’est pas le même pour tous les neurones.

Dans le cas des neurones amyéliniques, le potentiel d’action possède la même amplitude tout du long ; la dépolarisation en un point induit la dépolarisation du point voisin, la propagation est lente. Il existe une période réfractaire. Le potentiel d’action se déplace en sens unique du cône d’émergence vers les terminaisons.
Dans le cas des neurones myélinisés, il y a création successive des potentiels d’action le long de l’axone ; ceux-ci vont s’éloigner du site d’excitation initiale. La vitesse de transmission dépend du diamètre de la fibre (s’il augmente, la vitesse augmente). La conduction est dite conduction saltatoire (de nœud en nœud) et la propagation est rapide. Après la repolarisation, la membrane demeure inerte un certain temps ; les canaux à Na ne peuvent pas s’ouvrir (période réfractaire). Au niveau des nœuds de Ranvier on met en évidence une grande concentration de canaux sodique voltage dépendant qui sont responsable de la propagation du potentiel d’action.
La synapse correspond au point de connexion fonctionnel existant entre deux neurones. Un millimètre cube de substance grise du cortex peut contenir 5 millions de synapses. Les synapses peuvent être électriques ou chimiques :
Les synapses chimiques, quant à elles, sont présentent uniquement dans le tissu nerveux et ce sont elles que nous allons expliquer dans la suite de ce cours. Suivant les cellules impliquées on aura des :
Il peut être inhibiteur, on parle de potentiel post-synaptique inhibiteur (PPSI). Le PPSI est créé par l’entré d’ion chlorure Cl- ou la sortie d’ion potassique K qui permettent une hyperpolarisation diminuant l’excitabilité neuronale.
Le potentiel d’action est soumis à la loi du tout ou rien, c’est-à-dire que quelque soit le courant créé par le stimulus, s’il arrive jusqu’au seuil du neurone il permettra une dépolarisation complète, mais s’il n’arrive pas jusqu’au seuil, il ne se passera rien du tout :
synapse.
La période réfractaire est la période durant laquelle l’axone ne pourra plus propager de potentiel d’action après un premier potentiel d’action. Il y a présence de deux types de périodes réfractaires suivant l’avancé du premier potentiel d’action.
Le système nerveux végétatif (on autonome) est chargé de l’innervation du milieu intérieur, son champs d’innervation concerne les viscères, les glandes exocrines et endocrines et la vaso-motricité.
Au plan moteur, il innerve donc toutes les fibres musculaires lisses.
Au plan sensitif, il transmet la sensibilité viscérale, qui s’exprime par la sensation d’hyperpéristaltisme, la douleur par tension ou réplétion des viscères creux, par compression ou hypertrophie d’un viscère.
Le système nerveux végétatif intervient dans le contrôle et la régulation de la pression artérielle, la sécrétion et la mobilité gastro-intestinale, la vidange de la vessie, la sudation, la température corporelle, le rythme cardiaque et d’autres fonctions que nous verrons par la suite.
Dans ces régulations, le système nerveux végétatif joue certes un rôle actif mais il est lui-même sous le contrôle d’autres parties du système nerveux tel que l’hypothalamus, le tronc cérébral et certaines parties du système limbique.
Ces circuits sont souvent mis en jeu de façon réflexe, l’information acquise par des récepteurs viscéraux est transmise à des « centres » situés dans le tronc cérébral ou l’hypothalamus, la réponse appropriée est déclenchée au niveau de ces centres et transmise à la périphérie grâce aux fibres nerveuses effectrices du système nerveux autonome (SNA).
Une des caractéristiques du SNA est sa capacité à répondre rapidement aux sollicitations et changer ainsi par la suite les fonctions viscérales.
Par exemple, il peut doubler la fréquence cardiaque en moins de 3 à 5 secondes, doubler la pression artérielle en moins de 10 à 20 secondes.
À l’inverse la pression artérielle peut chuter en moins de 4 à 5 seconde et provoquer un évanouissement, la sudation peut apparaître en quelques secondes et la vessie se vider involontairement très rapidement.
II Organisation générale du systeme nerveuxautonome
II.A. Rappel anatomique sur le système nerveux
Le fonctionnement des organes du corps humain est soumis à une commande (ou régulation) assurée par un système de cellules spécialisées appelé le système nerveux.
Ce système peut être divisé en 2 grands systèmes:
• Le système nerveux cérébro-spinal affecté à la vie de relation qui assure les fonctions de motricité volontaire et de sensibilité.
Il est représenté par:
• Le système nerveux central (SNC) qui comprend le cerveau et la moelle épinière; et
• Le système nerveux périphérique (SNP) qui comprend les nerfs crâniens et les nerfs rachidiens (spinaux)
• Le système nerveux organo-végétatif appelé encore système nerveux autonome qui assure le contrôle et la régulation des fonctions dites végétatives.
Le fonctionnement du SNA se fait indépendamment de notre volonté et de notre conscience.
Il est divisé en 2 grands systèmes:
• Le système sympathique (orthosympathique) qui correspond à la région thoraco-lombaire et le système parasympathique qui comprend la portion crânienne et sacrée.
II.B. Le système nerveux sympathique
Les fibres sympathiques efférentes sont issues de la moelle épinière au niveau des segments T1(thoracique ou dorsale) jusqu’à L2 (lombaire).
Chaque fibre sympathique est composée de deux neurones; un neurone pré-ganglionnaire et un neurone post-ganglionnaire.
II.B.a. Neurone pré-ganglionnaire
Le corps cellulaire du neurone pré-ganglionnaire est situé dans la corne intermédio-iatérale de la moelle épinière et la fibre émerge par la racine antérieure de la moelle dans le nerf spinal correspondant.
Après son émergence le neurone sympathique pré-ganglionnaire quitte le nerf spinal par le rameau communicant blanc RCJ3) et pénètre dans le ganglion de la chaîne sympathique située de part et d’autre de la colonne vertébrale.
Le devenir du neurone pré-ganglionnaire est variable:
• Il fait synapse avec le neurone post-ganglionnaire dans le ganglion
• Il traverse le ganglion et se dirige vers le haut ou vers le bas de la chaîne pour faire relais dans un autre ganglion
• I1 peut traverser la chaîne para-vertébrale et cheminer vers un autre nerf sympathique innervant un ganglion pré-vertébral (ganglions coeliaque et hypogastrique) situé à distance de la chaîne sympathique para-vertébrale.
II.B.b. Neurone post-ganglionnaire
Le neurone post-ganglionnaire naît d’un ganglion para-vertébrale ou pré-vertébral et se dirige pour aller innerver les différents organes.
Certaines fibres post-ganglionnaires passent dans les nerfs rachidiens par l’intermédiaire des rameaux communicants gris (RCG).
Elles assurent l’innervation sympathique des vaisseaux sanguins, des glandes sudoripares et des muscles pilo-érecteurs (érecteurs des poils)
II.B.c. Distribution segmentaire des nerfs sympathiques
• Les fibres sympathiques du segment thoracique T1 se dirigent vers la tête
• celles du segment T2 vers le cou,
• celles des segments T3, T4, T5 et T6 vers le thorax,
• celles des segments T7, T8, T9, T10 et T 11 vers l’abdomen,
• celles des segments T12, Li et L2 vers les membres inférieurs.
Cette distribution est approximative car les chevauchements sont fréquents.
La distribution des neurones sympathiques vers les organes cibles dépend de l’origine embryonnaire des organes.
Le cœur par exemple reçoit de nombreuses fibres provenant de la chaîne sympathique du cou car le cœur se développe à partir du cou.
De même, les organes abdominaux reçoivent leur innervation des organes thoraciques car l’intestin primitif est originaire de cette région.
II.C. Système nerveux parasympathique
II.C.a. Distribution du système nerveux parasympathique
La structure anatomique du système parasympathique est très différente de celle du système sympathique.
Les fibres parasympathiques quittent le système nerveux central:
• En haut (au niveau bulbo-protubérentiel du tronc cérébral) par les paires crâniennes moteur oculaire commun (III), facial (VII), glosso-pharyngien (IX) et le vague ou pneumogastrique (X)
• En bas (au niveau sacré) par les 2ème, 3eme et 4ème nerfs sacrés.
Environ 75% des fibres nerveuses parasympathiques cheminent dans le nerf vague innervant les régions thoracique et abdominale du corps (coeur, poumons, l’estomac, l’intestin, la moitié du colon proximal, le foie, la vésicule biliaire, le pancréas et la partie haute des uretères).
• Les fibres parasympathiques du III innervent les sphincters pupillaires et les muscles ciliaires de l’oeil.
• Les fibres parasympathiques du VII innervent les glandes lacrymales, les glandes nasales et les glandes sub-mandibulaires.
• Les fibres du IX innervent la glande parotide (glande salivaire se trouvant dans le conduit auditif interne).
• Les fibres parasympathiques sacrées se rassemblent pour former les nerfs pelviens au niveau des plexus sacrés de chaque coté de la moelle.
Ces fibres se distribuent au côlon descendant, au rectum, à la vessie et à la partie basse des uretères et les organes génitaux externes (impliquées dans la stimulation sexuelle)
II.C.b. Les neurones parasympathiques pré- et post–ganglionnaires
L’innervation parasympathique est constituée de neurones pré- et post-ganglionnaire.
L’innervation parasympathique post-ganglionnaire de la tête provient des ganglions ciliaires (innervation de l’oeil), des ganglions ptérygo-palatin ou sphéno-palatin (innervation des glandes lacrymales, du nez et du pharynx), des ganglions sous-maxillaires (sub-mandibulaire) et optique (innervation des glandes salivaires et de la cavité buccale).
En revanche dans le reste du système parasympathique, le neurone pré-ganglionnaire atteint directement l’organe cible
Le ganglion et les fibres post-ganglionnaire sont situés dans l’organe; la synapse entre fibres pré- et post-ganglionnaires est située dans l’organe.
Les fibres post-ganglionnaires sont courtes de quelques millimètres à quelques centimètres (contrairement au système sympathique)
III Caractéristiques des fonctions sympatiques et parasymathiques
III.A. Les fibres adrénergiques et cholinergiques
Les fibres nerveuses sympathiques et parasympathiques secrètent l’un des deux neurotransmetteurs synaptiques; l’acétylcholine ou la noradrénaline.
• Les fibres libérant l’acétylcholine sont appelées fibres cholinergiques et
• Celles qui libèrent la noradrénaline sont nommées fibres adrénergiques.
Tous les neurones pré-ganglionnaires sont cholinergiques dans les systèmes sympathique et parasympathique.
Les neurones post-ganglionnaires du système parasympathique sont aussi cholinergiques.
La plupart des neurones post-ganglionnaires sympathiques sont adrénergiques à l’exception des neurones qui innervent les glandes sudoripares, les muscles pilo-érecteurs et de rares vaisseaux sanguins qui sont des neurones cholinergiques.
Les structures moléculaires de l’acétylcholine et de la noradrénaline sont :
• Acétylcholine
• Noradrénaline
III.B. Mécanismes de synthèse et de destruction de l’acétyLcholine et de La noradrénaline
III.B.a. L’acétylcholine
L’acétylcholine est synthétisée à partir de la choline dans le cytoplasme du corps cellulaire (cytosol).
La choline est acétylée par une choline-acétyltransférase avec la présence de l’acétyl-CoA pour donner l’acétylcholine, puis transférée et stockée à l’intérieur des vésicules des terminaisons nerveuses.
Elle est libérée sous l’action de l’influx nerveux avec la participation du calcium, mobilisé dans l’espace synaptique.
L’excès de neuromédiateur et la fraction liée aux récepteurs post-synaptique (nicotinique ou muscarinique) sont dégradées par l’acétylcholine-estérase qui libère l’acide acétique et la choline qui pourra être recaptée par le neurone.
III.B.b. Noradrénaline
La synthèse de la noradrénaline débute dans le cytosol et se termine à l’intérieur des vésicules.
La chronologie de la réaction de synthèse est:
Hydroxylation Tyrosine ——————- Dopa Décarboxylation
Dopa——————— Dopamine
Transport et stockage de la dopamine dans les vésicules
Hydroxylation – Dopamine———————- Noradrénaline
Dans la médullosurrénale, les réactions se poursuivent afin de transformer 8O% de la noradrénaline en adrénaline:
• méthylation Noradrénaline————— , Adrénaline
• La noradrénaline est stockée, avec la dopamine, par un mécanisme dépendant du calcium dans les vésicules en association avec l’ATP.
La libération se fait par l’arrivée de l’influx nerveux.
L’inactivation de la noradrénaline résulte:
• Principalement, de la diffusion dans les fluides de voisinage et vers la circulation sanguine
• De la recapture ; qui peut être de deux type,
• de type 1, neuronale, dans la terminaison pré-synaptique spécifique de la noradrénaline
• de type 2, extra neuronale, dans les structures post-synaptiques et le tissus périphériques, pour la noradrénaline comme pour l’adrénaline
• Du catabolisme enzymatique (transformation enzymatique) qui survient en 2 étapes:
• Dans la première interviennent
• la MonoAmine Oxydase (MAO) présente dans les terminaisons nerveuses adrénergiques
• La Cathécol-O-Méthyl Transférase (COMT) présente dans les terminaisons nerveuses et structures post-synaptiques (tissus)
• Dans la 2 étape interviennent,
Une aldéhyde déshydrogénase, de façon prédominante en péri et conduit à la formation d’acide vanylmandélique ou VMA
Une aldéhyde réductase, de façon prédominante dans le système nerveux central, et conduit à la formation de MOPEG (3-métl-4-hydroxy-phényléthylène-glycol)
III.C. Les récepteurs de ces neuromédiateurs
Avant que l’acétylcholine, la noradrénaline ou l’adrénaline provoquent leur effet sur l’organe effecteur, ces médiateurs doivent se fixer sur leurs récepteurs spécifiques situés sur la membrane post-synaptique, la fixation du neuromédiateur sur le récepteur produit une modification moléculaire de la protéine du récepteur provoquant soit une ex soit une inhibition de la cellule cible.
III.C.a. Les récepteurs cholinergiques
Les récepteurs à l’acétylcholine sont de deux types ; nicotinique et muscarinique.
1 Récepteurs muscariniques
Appelés ainsi parce qu’ils sont stimulés spécifiquement par la muscarine.
Il existe 4 sous-types de ces récepteurs:
• M1 post-synaptique ganglionnaire (rôle min et sur organes cibles)
• M2, M3, M4 se localisent sur les organes i par les fibres du système parasympathique et les fibres cholinergique du système sympathique.
2 Récepteurs nicotiniques
Appelés ainsi parce que stimulés spécifiquement par la nicotine.
Ces récepteurs sont essentiellement situés au niveau des relais ganglionnaires entre le premier neurone et le deuxième neurone (sous type N1) tant sur la voie sympathique que sur la voie parasympathique
Un second sous-type N2 se distribue au niveau des jonctions neuro-musculaires de la plaque motrice des muscles striés.
III.C.b. Les récepteurs adrénergiques
Ils sont classés en 5 sous-types principaux:
1 Récepteurs alpha
1 Sous-type Alpha 1
Post-synaptique, produisant essentiellement une stimulation.
2 Sous-type Alpha
Pré-synaptique, inhibant la libération de la noradrénaline, quelques récepteurs alpha 2 sont post-synaptiques dans la système nerveux central et sont inhibiteurs du tonus sympathique.
2 Récepteurs bêta
1 Sous-type Bêta I
Post-synaptique, produisant une stimulation.
2 Sous-type Bêta 2
Post-synaptique, à effet inhibiteur (relaxation)
Le récepteur bêta pré-synaptique situé sur les terminaisons synaptiques ont un rôle facilitateur sur l’activité synaptique (leur blocage par les bêta bloquants réduit l’activité sympathique, ce qui expliquerait l’effet de relâchement artériel de ces substances dans les traitements chroniques)
3 Sous-type Bêta 3
Post-synaptique, plus récemment mis en évidence, produisant une lipolyse dans les adipocytes.
IV Les effets de la stimulation nerveuse végetative
Les effets de la stimulation des systèmes sympathique ou parasympathique dépendent de l’organe concerné.
La stimulation d’un système peut entraîner des effets excitateurs sur certains organes mais inhibiteurs sur d’autres.
IV.A. Œil
IV.A.a. La pupille
La stimulation sympathique entraîne la contraction des fibres radiaires de l’iris et dilate donc la pupille.
La stimulation parasympathique contracte le muscle circulaire de l’iris et rétrécie donc la pupille.
Le parasympathique est stimulé de façon réflexe lorsqu’une lumière intense pénètre dans l’oeil; ce qui réduit l’ouverture de la pupille et diminue la quantité de lumière qui atteint la rétine.
IV.A.b. Le cristallin
La convergence de la lentille est entièrement contrôlée par le parasympathique.
La lentille est maintenue aplatie grâce aux ligaments radiaires.
L’excitation parasympathique contracte le muscle ciliaire libérant la tension des ligaments du cristallin et lui permet de devenir plus convexe, ce qui fait converger le regard sur un objet proche.
IV.B. Les glandes
Les glandes nasales, lacrymales, salivaires et les glandes de la partie haute du tube digestif sont stimulées par le parasympathique qui provoque une hypersécrétion.
Les glandes sudoripares sont essentiellement innervées par le sympathique qui provoque une sécrétion abondante.
Les glandes apocrines dans les aisselles (la même origine embryologique que les glandes sudoripares).
Elles dépendent uniquement du sympathique, elles synthétisent des sécrétions épaisses et odoriférantes.
IV.C. Le tube digestif
Le tube digestif possède son propre réseau nerveux intrinsèque appelé plexus intramural.
La stimulation parasympathique augmente le péristaltisme et relâche les sphincters, accélérant ainsi la progression du contenu alimentaire du tube digestif.
La stimulation sympathique a des effets opposés ; inhibition du péristaltisme et contraction des sphincters, donc ralentissement du bol alimentaire.
IV.D. Le cœur
La stimulation sympathique augmente le rythme cardiaque et augmente la force de contraction.
La stimulation parasympathique entraîne un ralentissement du rythme.
IV.E. Les vaisseaux sanguins
La stimulation sympathique contracte la plupart des vaisseaux (vasoconstriction), particulièrement au niveau de la peau et des viscères abdominaux.
La stimulation parasympathique est sans effet, sauf au niveau due la face où elle les dilate (lorsque le visage rougit)
IV.F. La pression artérielle
La stimulation sympathique, par l’intermédiaire de son effet dur le coeur, a tendance à augmenter la pression artérielle. Le parasympathique a un effet opposé.
IV.G. La vessie
L’effet majeur de l’innervation sympathique de la vessie est d’empêcher la vidange lorsque celle-ci n’est pas pleine (inhibition sur le détrusor et excitation sur le trigone et le sphincter interne)
L’effet du parasympathique entraîne une miction (contraction du détrusor et relâchement du trigone et du sphincter)
IV.H. L’Appareil génital mâle
La stimulation sympathique a un effet positif et entraîne une éjaculation.
La stimulation parasympathique a un effet positif et entraîne une érection.
IV.I. La peau
La stimulation sympathique entraîne une piloérection (redressement du poil)
IV.J. La médullosurrénale
La médullosurrénale reçoit une innervation sympathique comportant une voie effectrice à un seul neurone, la cellule médullosurrénalienne se substitue au second neurone et assure la «médiation adrénergique» qui prend ici un caractère humoral.
La stimulation nerveuse sympathique de la médullosurrénale entraîne une libération importante d’adrénaline et de noradrénaline dans la circulation sanguine.
Ces deux médiateurs ont les mêmes effets sur les différents organes que la stimulation sympathique, sauf que leur durée d’action (de ces hormones) est 5 à 10 fois plus longue puisque les catécholamines sont dégradées lentement dans le sang.
V En conclusion
On peut dire que le système parasympathique est un système de repos qui est destiné à conserver les ressources de l’organisme et sa nutrition (sécrétion salivaire, gastro-intestinale, contractilité du sphincter pupillaire, l’accommodation à la vision de près, inhibition des fonctions cardiaques…)
Tandis que le système sympathique est au contraire un système réagissant à toute situation critique (exercice musculaire violent, la douleur, le froid, la peur, la colère, l’angoisse, le stress…) par une mise en état de défense de l’organisme appelé une décharge massive.
Dans ces situations la pression artérielle, le débit sanguin, la glycémie, et la glycolyse, la force musculaire et l’activité mentale augmentent et le métabolisme cellulaire est accru.
